Photosynthesis
Photosynthesis was first described in the 19th century as the process by which plants and some bacteria and algae trap light energy (radiant energy / electromagnetic radiation either from the sun or from other light sources) and use the energy to synthesize glucose which is then used to synthesize other carbohydrates such as fructose (in fruits), sucrose (in sugar cane and sugar beets), starches (potatoes, rice and other grains), cellulose (mainly used to build cell walls which as essential in giving plant stems rigidity and provide a support structure for flowers and fruits).
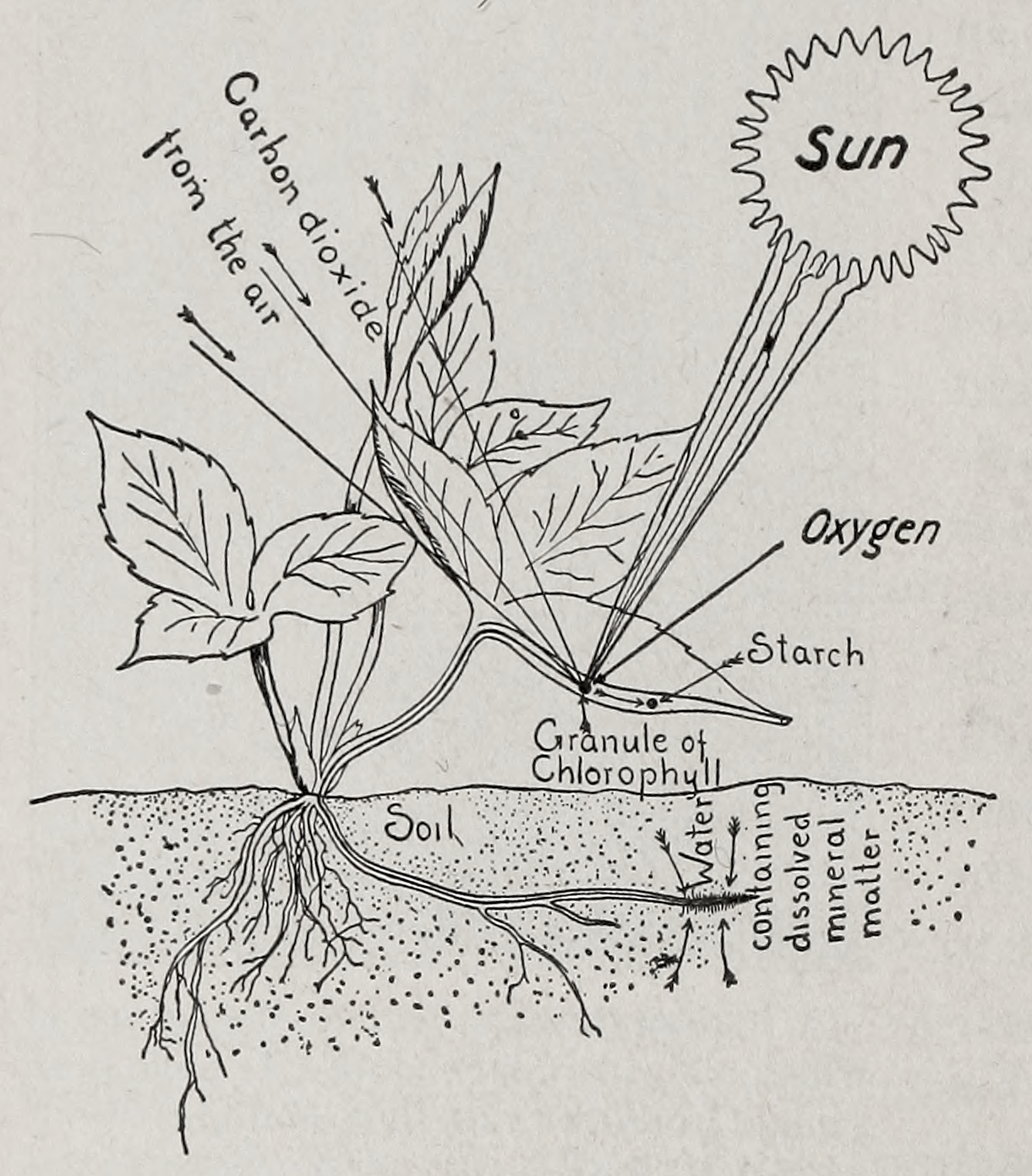
Image: A schematic representation of photosynthesis. (Hunter, 1928).
Most photosynthetic organisms utilize carbon dioxide as the source of carbon, and water, as the source of hydrogen and oxygen. Some photosynthetic organisms use organic carbon as the source of carbon. Oxygen is released as a by-product in some organisms while other organisms do not produce oxygen. Therefore, photosynthetic organisms can be classified as oxygenic (the majority) or anoxygenic. The general formula for photosynthesis is:
In plants and algae, photosynthesis takes place in organelles called chloroplasts. A typical plant cell contains about 10 to 100 chloroplasts. Plants absorb light using a pigment called chlorophyll, the pigment that gives plants their green color. Along with the green pigment are yellow to orange pigments called carotenes and xanthophyll pigments respectively. Most of the year these colors are masked by the great amounts of the green pigment. Algae also use chlorophyll, but various other pigments are present such as phycocyanin, carotenes, and xanthophylls in green algae, phycoerythrin in red algae (rhodophytes) and fucoxanthin in brown algae. In the fall, due to changes in daylight and temperature, the leaves stop the photosynthesis process. The chlorophyll breaks down, the green color disappears, and the yellow to orange colors become visible and give the leaves the fall colors we observe.
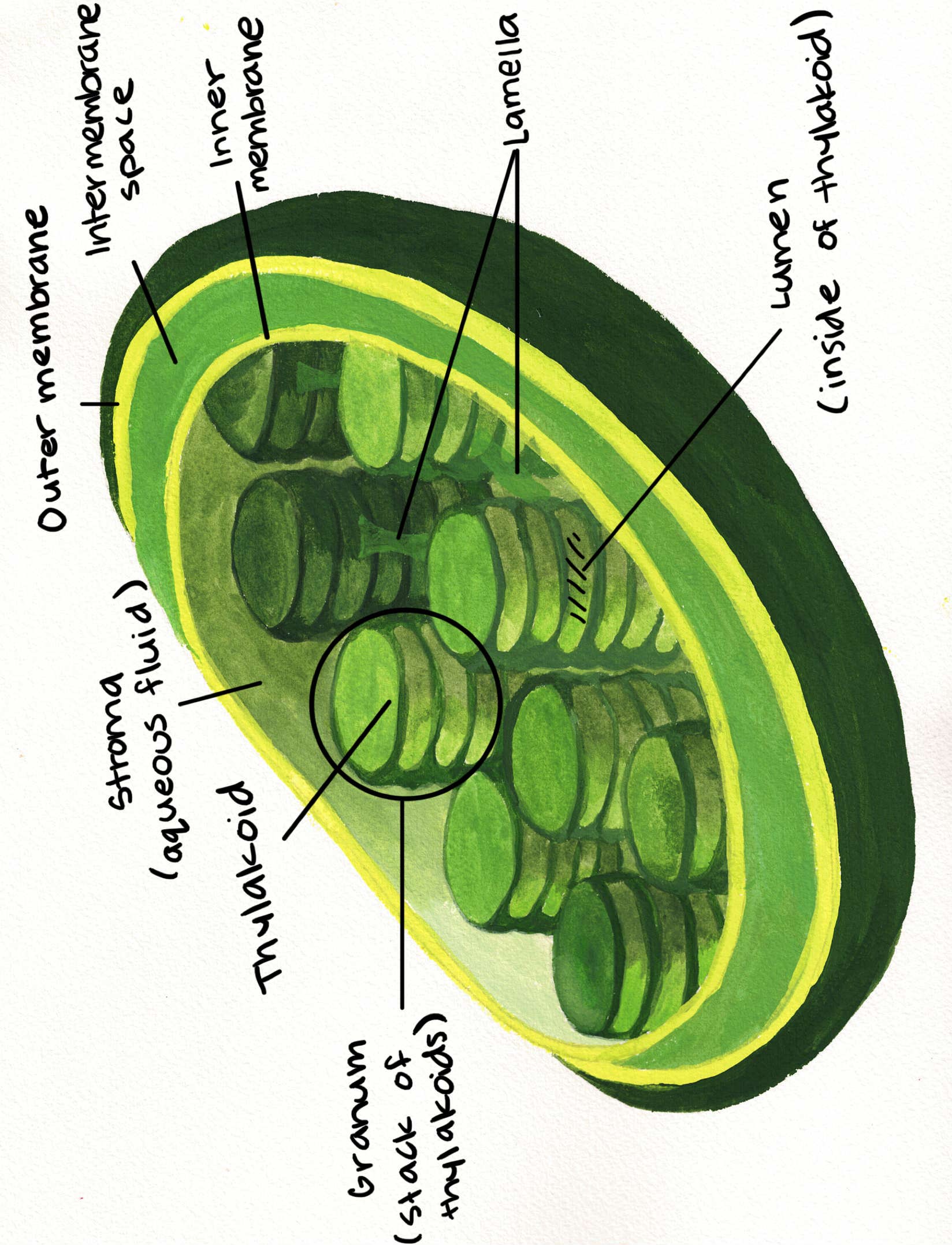
Parts of the chloroplast:
Left: Distribution of chloroplasts inside several plant cells. (Source CC BY-SA 3.0, No alterations made).
Right: The structure of a chloroplast showing thylakoids, grana and the stroma.
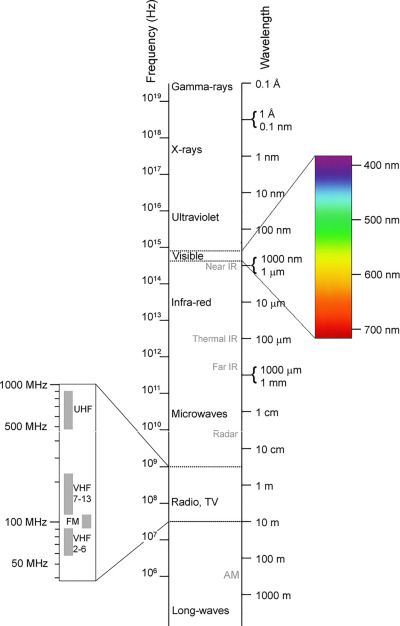
The electromagnetic spectrum. (Source: Wikipedia, CC BY-SA 3.0, No alterations made)
White light from the sun is composed of several individual colors which differ in their wavelengths resulting in the electromagnetic spectrum. While chlorophyll molecules absorb red and blue light efficiently, these molecules reflect green light, thereby giving them a green color.
There are two types of chlorophyll pigments: chlorophyll a, which absorbs violet-blue and orange-red, and chlorophyll b, which absorbs blue and yellow.
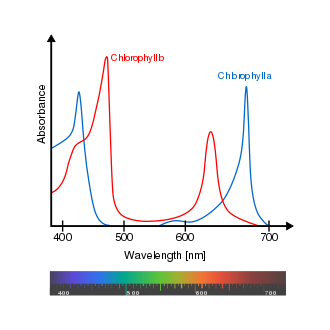
Light wavelength absorbance for chlorophyll a and b. (Source: Wikipedia, CC BY-SA 3.0, No alterations made)
Pigments are organized into clusters called Photosystems or Light harvesting complex . There are two types of Photosystems, I and II. Generally, photosystems are transmembrane protein complexes located in the thylakoid membrane.
Photosynthesis occurs in two steps;
This step occurs on the thylakoid membrane. Light is absorbed by the photosystem II (PSII), this light energy is used for two main functions, the first is to split water into H, O and releases electrons, this process is called Photolysis. The electrons are used to pump H+ into the thylakoid lumen (inside). The second effect of the light energy is to cause excitation of electrons at the PSII. The excited electrons are transferred to plastoquinone, which then passes them to the cytochrome complex. The process of electrons moving from the high energy / excitatory phase to low energy results in the movement of protons (hydrogen ions) into the thylakoid lumen. Electrons that are now in a reduced energy state are transferred to the PSI. This means there is now more hydrogen ions in the thylakoid lumen than in the stroma. This proton gradient (together with the proton gradient produced by the water-splitting complex in PS II) is used to make ATP via ATP synthase. When the electrons are delivered to the PSI, light again strikes this photosystem and electrons gain energy again and move higher in their excitatory state. The electrons join up with NADP+ and H+ and reduces it to NADPH. ATP (formed by the ATP synthase) and NADPH go to the light independent reaction in the stroma.
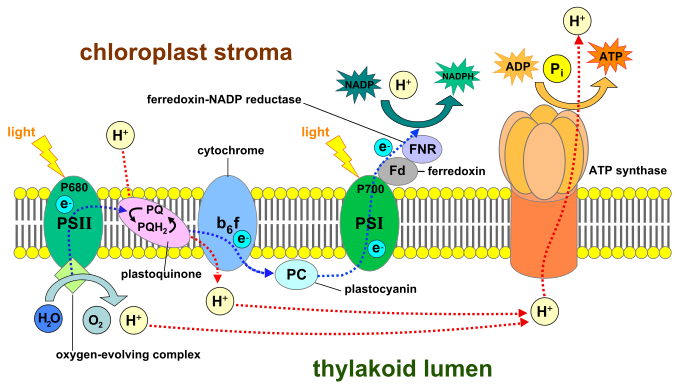
An illustration of the light dependent reaction. (Source: Wikipedia, CC BY-SA 3.0, No alterations made)
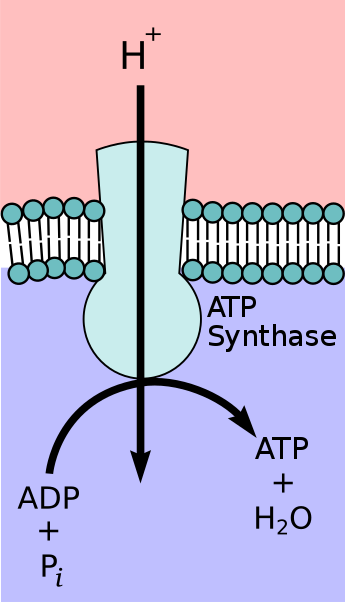
An illustration of how the ATP synthase membrane protein catalyzes the formation of ATP using the movement of hydrogen ions. (Source: Wikipedia, CC BY-SA 3.0, No alterations made)
This will be reviewed again in Cellular respiration, so briefly, ATP synthase is an enzyme located across the thylakoid membrane. Movement of Hydrogen ions through it makes the enzyme rotate and brings ADP together with a phosphate group resulting in the formation of ATP.
The light independent reaction is also called the Calvin cycle (developed by Melvin Calvin). This step utilizes the H and NADPH from the light dependent reaction. ATP, generated in the light dependent reaction is used to power the Calvin cycle.
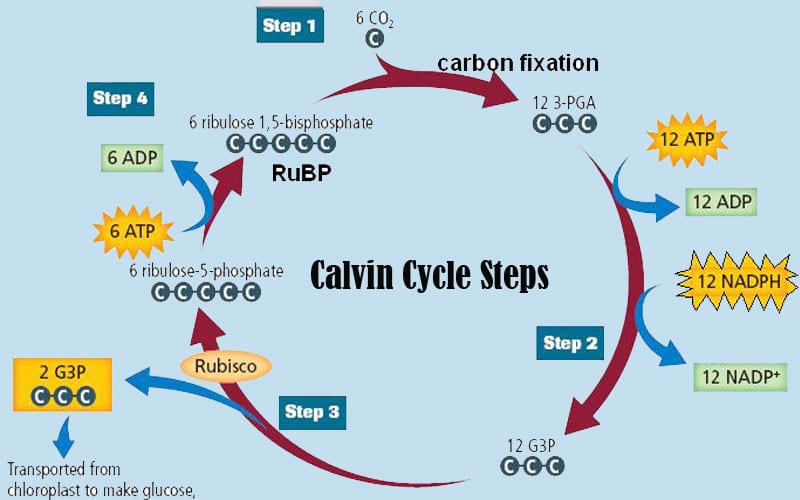
A detailed illustration of the Calvin cycle. (Source: Wikipedia, CC BY-SA 3.0, No alterations made)
The first reaction involves fixing Carbon dioxide by Ribulose Bisphosphate (RuBP). RuBP is a 5 carbon sugar, when CO2 is added it becomes a 6 Carbon sugar but this sugar is very unstable so it immediately splits into 2 3C sugars called Phosphoglyceric Acid (PGA).
PGA uses ATP to remove the hydrogen from NADPH to form a new molecule called Phosphoglyceraldehyde (PGAL), which is still a 3-carbon molecule. Every 6 Calvin cycles result in 12 PGAL molecules. 2 of these are used to produce a glucose molecule and the remaining 10 are recycled to make more RuBP. PGAL can also be used as an energy source directly, or can be used to produce fat and/or proteins.
Fates of PGAL
Many photosynthetic organisms have not developed mechanisms to concentrate carbon dioxide, which would increase the CO2 levels available to the initial carboxylase of the Calvin cycle, the enzyme RuBisCO. The benefits of such mechanisms include increased tolerance to low concentrations of inorganic carbon, and reduced losses through photorespiration. As a result, carbon dioxide concentrating mechanisms can make plants more tolerant to heat and water stress.
CAM Plants
CAM plants use the Crassulacean acid metabolism, also known as CAM photosynthesis, as an adaptation to arid conditions. CAM metabolism is a carbon fixation pathway that evolved in some plants to photosynthesize during the day, but only exchange gases at night. In plants that utilize CAM fully, the stomata in the leaves remain shut during the day to reduce evapotranspiration, but they open at night to collect carbon dioxide and allow it to diffuse into the mesophyll cells. Sixteen thousand species of plants, including the cactus, use CAM photosynthesis.
C4 Plants
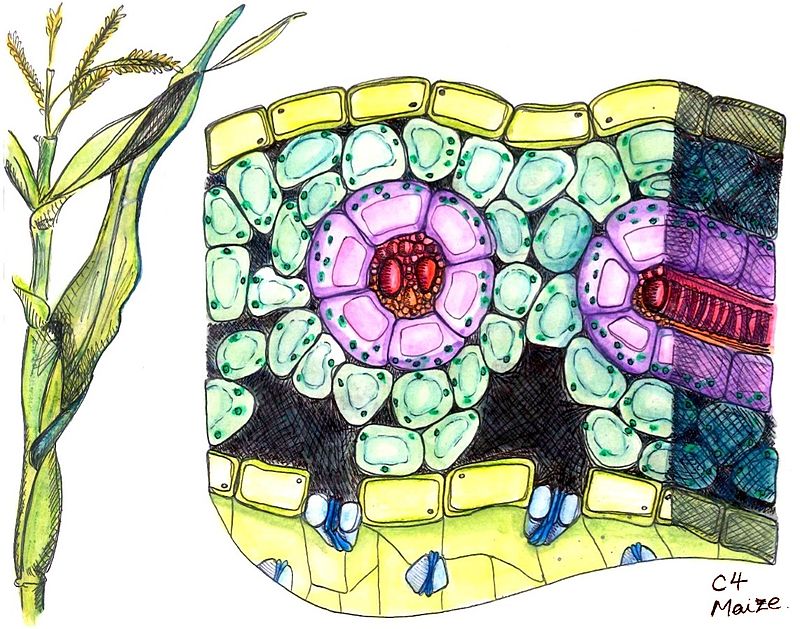
C4 plants incorporate CO2 into one of the 4-carbon compounds, malic acid or aspartic acid. C4 plants have a distinctive internal leaf anatomy called kranz anatomy. See image alongside. Their vascular bundles are surrounded by two rings of cells; the inner ring, called bundle sheath cells, contains starch-rich chloroplasts lacking grana, which differ from those in mesophyll cells present as the outer ring. Hence, these chloroplasts are called dimorphic. The primary function of kranz anatomy is to provide a site in which CO2 can be concentrated around RuBisCO, thereby avoiding photorespiration. Mesophyll and bundle sheath cells are connected through numerous cytoplasmic sleeves called plasmodesmata whose permeability at leaf level is called bundle sheath conductance. A layer of suberin is deposed at the level of the middle lamella to reduce the apoplastic diffusion of CO2 (i.e., leakage). Examples of C4 plants include tropical grasses, such as sugar cane and maize, many broadleaf plants tare also C4 plants. About 7600 species of terrestrial plants use C4 carbon fixation.
C3 plants
The large majority of plants are C3 plants because, as described previously, the carboxylation products of the Calvin cycle are 3-carbon compounds. They lack C4 dicarboxylic acid cycles, and therefore have higher CO2 compensation points than CAM or C4 plants.
You can access Tensai High School Biology revision questions HERE